
-
![画像]()
フェミウスイナズマ複合種群。左上:フェミウスイナズマ♂、左下:フェミウスイナズマ♀、中上:イポナイナズマ♂、中央下:イポナイナズマ♀、右上: ユーフェミアイナズマ♂、右下:フェミウスイナズマとイポナイナズマの中間的個体♀
-
![画像]()
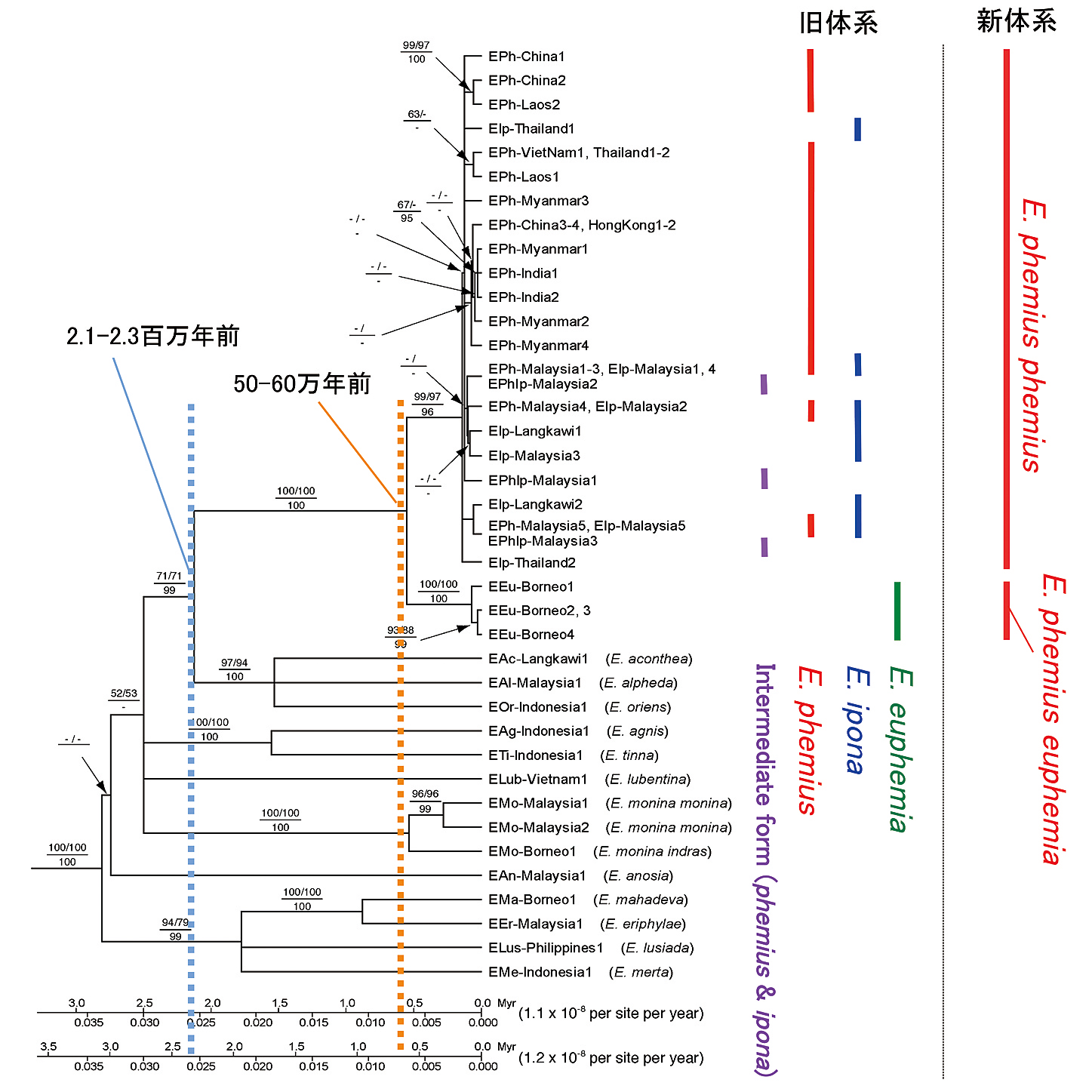
フェミウスイナズマ複合種群の分子系統樹。Yago et al. (2012)を改変、樹形はミトコンドリアDNAに基づいて近隣結合法により構築。 各枝の数値は近隣結合法(左上)と最節約法(右上)によるブートストラップ値、ベイズ法(下)による事後確率で、ブートストラップ値50 %以上と事後確率0.9以上が示されている
-
![画像]()
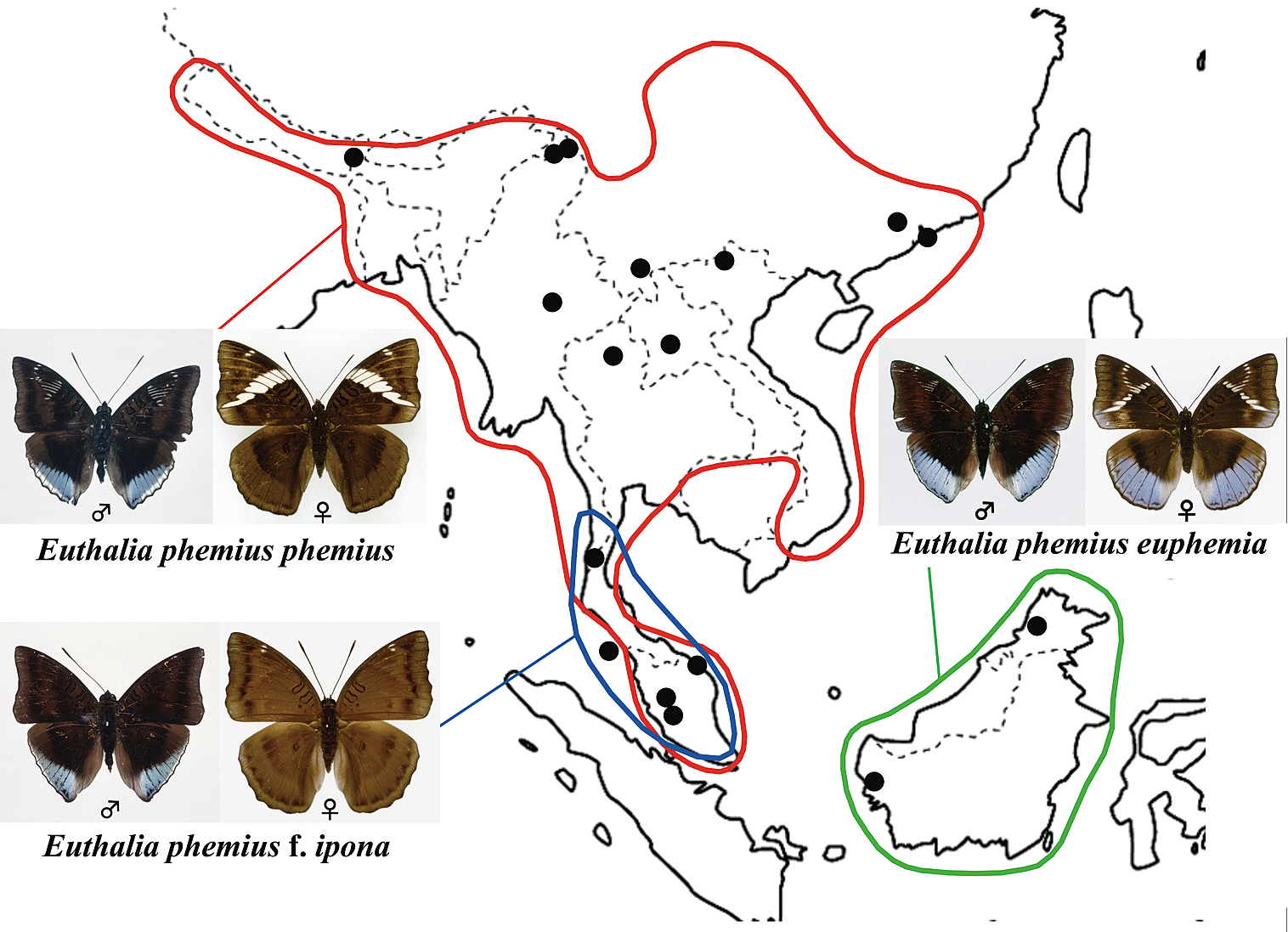
フェミウスイナズマ複合種群の地理的分布。Yago et al. (2012)を改変。赤枠:フェミウスイナズマ、青枠:イポナイナズマ、緑枠:ユーフェミアイナズマ、黒丸は研究材料に用いた標本の産地を示す。翅の斑紋による違いから、かつてはそれぞれ別種として見なされていたが,現在ではすべて同一種として扱われている

B24
フェミウスイナズマ複合種群
翅の斑紋に惑わされた種分類
東南アジアに生息するタテハチョウ科・イナズマチョウ属Euthaliaのうち、中国南部からインド、インドシナ、マレー半島と広域に分布する前翅に白斑を持ったフェミウスイナズマEuthalia phemius、マレー半島に産する前翅に白斑のないイポナイナズマEuthalia ipona、ボルネオ特産で前翅に白斑かつ♀後翅に青帯があるユーフェミアイナズマEuthalia euphemiaの3種は、互いに形態が酷似することからフェミウス複合種群(Euthalia phemius complex)とも呼ばれ、上記のような斑紋の違いからそれぞれ別種として扱われていた。
ところが、核とミトコンドリア双方のDNAから分子系統解析を行ったところ、各種間の遺伝的差異はほとんど検出されなかった。次に種の分類として有効な♂交尾器の形態(特に♀を把握するvalvaの形状)をフーリエ変換という手法を用いて解析したところ、明確な差異を見出せなかった。その後の標本調査で、種分類の根拠となっていた斑紋に関しても、フェミウスイナズマとイポナイナズマとの中間的な斑紋を持つ個体がマレー半島で複数頭見つかった。
結果として、これら3種は同一種フェミウスイナズマにまとめるのが妥当とする結論に至り、ユーフェミアイナズマは本種の亜種に、イポナイナズマは本種の一型に変更された。かつて分子系統学的研究と伝統的な形態を主体とする分類学的研究とはデータが合わないこともあり、議論が対立していた時期もあったが、最近の研究の進歩により、分子系統解析は分類学的研究に極めて有効なツールとして今では認識されている。
一方、フェミウスイナズマの種・亜種の分化や分布形成過程も解明するため、分子データから分岐年代推定を行ったところ、本種の共通祖先はちょうど氷期と間氷期の激しい気候変動の繰り返しが世界中で見られるようになった時期にほぼ当たる2.1~2.3百万年前に出現し、その後、50~60万年前にマラッカ海峡の形成に伴う地理的分断により、マレー半島を含む大陸の集団とボルネオの集団に分岐して、それぞれの集団が各亜種に分化していったことが明らかにされている。 (矢後勝也)
参考文献 References
塚田悦造(編)(1991)『図鑑 東南アジア島嶼の蝶 第5巻 タテハチョウ編(下)』プラパック(松本)。
Yago, M. et al. (2012) Revision of the Euthalia phemius complex (Lepidoptera: Nymphalidae) based on morphology and molecular analyses. Zoological Journal of the Linnean Society 164: 304–327.